
|
|

Motion in Calmodulin [cm]
[ jump to morphs ]
Classification Known Domain Motion, Hinge Mechanism [D-h-2]
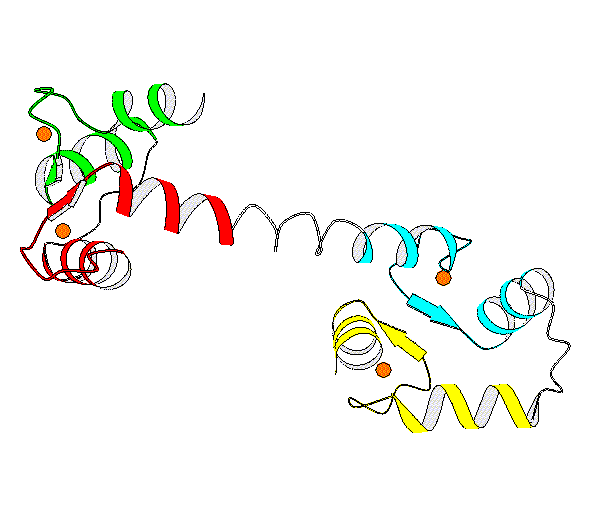
Structures 1CDL Closed (mammelian, recomb., X-ray) [ PartsList ] 1CLL Open (human, X-ray, refined) [ PartsList ] 1CTR Closed [ PartsList ] 2BBM fly, NMR, closed with peptideClosed [ PartsList ] 2BBN fly, NMR, closed with 2nd peptideClosed conf. 3 [ PartsList ] 4CLN fly, X-rayOpen [ PartsList ]
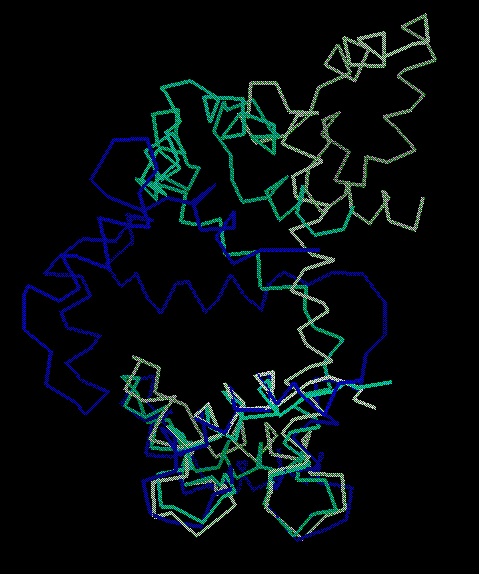
Description Basically, this hinge motion involves a long helix splitting into 2 helices (inclined at ~100 degrees) with a strand in between. The unligated form of calmodulin contains two globular domains, connected by a long helix. NMR and X-ray structures of ligated calmodulin show the molecule binding to peptide helices with different sequences and the two domains closing around the peptide far enough to make contact with each other. In this motion, the long interdomain helix, which is known to have only marginal stability in solution, partly unfolds to break into two helical segments connected by a 4-residue hinge region in an extended conformation. The angle between the axes of the two helical segments is ~100 degrees. As there is an additional twist around the helix axes, the total rotation of one domain relative to the other is upwards of 150 degrees. Calmodulin can bind peptides with different sequences because of flexibility in the side chains that make contact with the peptide and by slightly shifting the relative placement of the domains through changes in the extent of the hinge region, which has consequently been dubbed a variable 'expansion joint.' Specifically, in comparing 4cln with 2bbm, the largest torsion angle differences are in residues 72 to 82, where the protein changes from an alpha helix to an extended structure. This contains at most 20 changing torsions.
Particular values describing motion Annotation Level (1..10) = 7 Domain 1 (residue selection) = 2-80 Domain 2 (residue selection) = 81-147 Location of a Hinge (residue selection) = 72 - 82 (4cln v. 2bbm) Maximum CA displacement (A) = 60 (After sieve-fitting on domain-1) Maximum Rotation (degrees) = 148.02 Experimental Methods = xn (Traditional X-ray and NMR) Number of Inter-domain connections = 1 Number of hinges = 1 Number of Significant Torsion Angle Changes = 18 (Greater than 20 degrees) Creation Date = 19970822 Modification Date = 19970822
References L Stryer (1995). Biochemistry. New York, W H Freeman and Company. [Medline info for 11455603] M Ikura, G M Clore, A M Gronenborn, G Zhu, C B Klee and A Bax (1992). Solution structureof a Calmodulin-Target peptide complex by multidimensional NMR. Science. 256: 632-644. [Medline info for 92263094] W E Meador, A R Means and F A Quiocho (1992). Target enzyme recognition by Calmodulin: 2.4 Å structure of a Calmodulin-Peptide Complex. Science. 257: 1251-1255. [Medline info for 92390716] W E Meador, A R Means and F A Quiocho (1993). Modulation of calmodulin plasticity in molecular recognition on the basis of x-ray structures. Science. 262: 1718-1721. [Medline info for 94082290]
Data and Graphics Closed Form Adapted from Biochemistry, Copyright 1995, Lubert Stryer. Open Form Adapted from Biochemistry, Copyright 1995, Lubert Stryer. Torsion Changes + Atom Deviations Columns are, respectively: residue, phi-O, psi-O, sidechain-rotamer-O, phi-C, psi-C, rotamer-C, dphi, dpsi, dCA (after doing a fit) (O=open-form, C=closed-form). The closing of calmodulin in 3 steps From another viewpoint, showing open, closed, and partially open. 4x4 Transformation Matrix The following 4x4 matrix [1 .. 16] orients the opened form so that the axis of rotation is along the z-axis and the origin is at the molecular centroid.
GO terms associated with structures Molecular function protein serine/threonine kinase activity, ATP binding, calcium ion binding, protein kinase activity Biological process protein amino acid phosphorylation
Morphs
[ show all images ]
Best representative Morph Morph name Structure #1 Structure #2 Residues [ ] [ ]
![[help]](http://www0.molmovdb.org/images/help-icon.gif)
![[home]](http://www0.molmovdb.org/images/home-icon.gif)
![[movies]](http://www0.molmovdb.org/images/icon.movie.gif)
Copyright 1995-2005 M. Gerstein, W. Krebs, S. Flores, N. Echols, and others
Email: Mark.Gerstein _at_ yale.edu
/sicon.png)
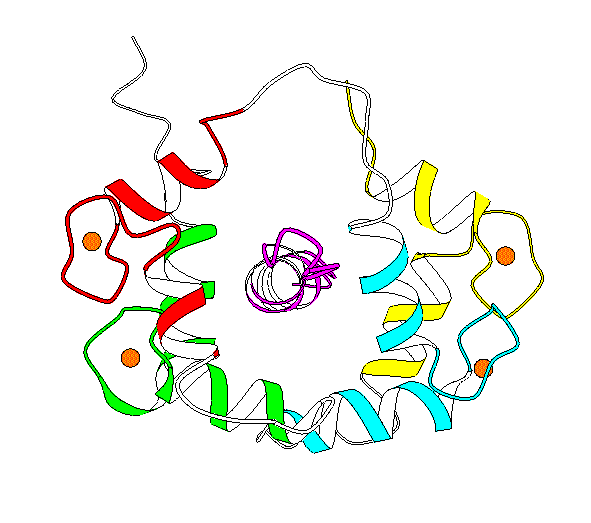{kind=link}
{kind=link}
{kind=link}